Extremophilic bacteria thrive in extreme conditions. Bacteria in deep sea vents have to withstand high heat and/or high pressure conditions. They must have adapted to their extreme environment by having specific genes that encode more resilient proteins or have special metabolic pathways. What many researchers have done is analyze the genome of extremophilic bacteria, then compare their unique proteins with proteins from microbes that live in more ordinary environments.
 |
| A deep sea vent, known as a black smoker, where diverse extremophilic microbes thrive. |
Bacteria from the deep sea were compared with shallow water counterparts by Dr. Lauro, such as Enterococcus faecalis with Carnobacterium. Dr. Reytor compared protein synthesis between extremophilic bacteria and temperate mesophiles such as common pathogens. Protein synthesis at these extreme conditions is made possible by deep sea extremophiles and other bacteria in extreme environments survive because their genome has additional genetic clusters, though their function is unknown. Dr. Costa, meanwhile, spent most of his time overviewing the metabolic pathways of these extremophiles, indirectly by measuring the concentrations of nitrite, nitrate, ammonia, and sulfide, as well as other organic and inorganic compounds within the native waters of the bacterium species. Costa's emphasis on the geochemistry and the setting of the bacteria will show where some thermophilic and extremophilic bacteria grow best.
Carnobacterium is found deep in the Aleutian Trench, yet is genetically similar to Enterococcus faecalis, suggesting that minimal evolution was needed to make the switch. For example, P. profundum strain SS9 and strain 3TCK have different bathytypes but show great synteny when their genome was sequenced (Lauro 20).
 |
| E. faecalis (pictured above) and Carnobacterium are genetically similar! |
“An essential cellular process inhibited by hydrostatic pressure is protein synthesis. Hydrostatic pressure induces the synthesis of a number of ribosomal and heat-shock proteins in Lactobacillus and Escherichia Coli.” However, many bacteria found in the vents are the exact opposite, having “an SOS response, with heat-shock genes being over-expressed at atmospheric pressure due to partial loss-of-function of the ribosomal units to create folded proteins” (Lauro 19). The most major difference genetically speaking between deep sea bacteria and shallow water ones were genes that coded for flagella synthesis and motility, as these are the most affected by pressure. One difference that was found was that deep sea vent bacteria had greater intergenic spacers. Additionally, a greater percent of rRNA copies were found on deep bathytypes than shallow water bacteria. Many genes were found in the genome of deep bathytypes coded for cell membrane unsaturation, as well as photolyase (Lauro 20). Photolyase repairs the cyclobutane prymidine dimers caused when blue light strikes DNA. When bacteria are this deep in the water, this gene is not necessarily needed to be expressed.
Lipases from Caldanaerobacter subterraneus subspecies tengcongenesis, and Thermoanaerobacter thermohydrosulfuricus were cloned and expressed in Escherichia coli. Lipase hydrolysizes and synthesizes long-chain acylglycerols, with a wide range of use in dairy, pharmaceutical, and the food industry in general. For instance, “the lipase from Thermoanaerobacter thermohydrosulfuricus were extremely S-stereoselective towards esters of secondary alcohols (Royter 769).” This is useful because it will create extremely pure compounds only. These lipases are extremely resistant to high temperature and were active across a wide range of pH. This is why lipase from extremophilic bacteria are so useful, because they can survive above 70 and 80 degrees. These enzymes are called “extremozymes.” These extremozymes are better studied when recombined with mesophilic bacteria, so they have been cloned and expressed in Escherichia coli, instead of their extremophilic microbe cousins.
Thermoanaerobacter thermohydrosulfuricus was isolated from Solar Lake, while Caldanaerobacter subterraneus subspecies tengcongenesis was taken from a Chinese hot spring. To make them reproduce, the agar they chose had to have special nutrients, specifically metal halides (Royter 771). They were incubated at 65 degrees, almost twice the optimal growth temperature of a typical mesophilic bacteria. Escherichia coli was cultivated in Luria-Bertani agar, with tryptone and 1% sodium chloride. Escherichia coli was transformed using heat-shock, and the DNA fragments that were cut up by HindIII and BamHI were analyzed with the PCR technique (Royter 771).
To determine what the lipase used as active sites, various chemical compounds were added into the culture. If the compounds inhibited the enzyme, that means the compounds either directly or indirectly competed with the normal substrate. Therefore, the active site must contain some of these groups. A spectroscope was used to see the effects of metal halides and other chemical compounds on purified lipase. Lipase was added with these compounds, then after a short amount of time, residual activity of the enzyme was measured (Royter 772). (Royter 777). “Serine and thiol groups are part of the active site for these enzymes (Royter 769).” As a result of this research, it was concluded that due to their active sites, the lipase and other enzymes of this nature came from mesophilic and psychrotrophic bacteria.
To determine the optimum pH for the operation of the lipase that was synthesized with cloned genes, turbidity was measured. Samples of lipase were faced to see how much copper soap could be used to create fatty unsaturated acids. Also, samples of the lipase with the lipase were subjected to acetic acid and sodium hydroxide for 70+ minutes (Royer 772). This questioned whether or not the lipases were stable with strong bases.
Also, the enzyme was subjected to two cycles of thawing and freezing. Some reduction was reported, but it was fairly resistant, with 70-80% remaining (Royter 775).
The enzyme was then placed within different solutions of esters to see the relative rate of hydrolysis. The enzymes were most effective with secondary alcohols (Royter 775).
After the transformation of Escherichia coli, lipase activity was measured in a supernatant without cells, but with cell lyase and para-nitrophenyl palmitate. This cell-free supernatant was created by precipitating heat, hydrophobic interaction, and then gel-filtering the substance. Residual activity is what it is called freed from the cell.
The results and conclusions were that over 17 amino acids were found to be similar in the terminal amino acid of the lipases between Thermoanaerobacter thermohydrosulfuricus and Caldanaerobacter subterraneus subspecies tengcongenesis. The genes were then sent through PCR and amplified. The optimum temperature for the lipase was 75 degrees, with a pH of about 8.0 (Royter 775). They remain active up to 90 degrees, as well. A 10% decrease in Thermoanaerobacter thermohydrosulfuricus lipase activity was measured when it was incubated at 85 degrees Celsius for 50 minutes. However, Caldanaerobacter subterraneus subspecies tengcongenesis saw an 80% reduction in residual lipase activity after the same incubation. These enzymes has an activity peak similar to the peaks with regular enzymes, with it skewing towards the right, with a sharp decline past the optimal temperature as the enzyme is denatured.
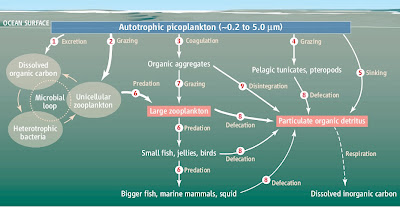 |
| Microbial loops play an important role in marine ecosystems, digesting marine snow. (click to enlarge!) |
Extremophiles are important because they represent more than half of all sea microbes live deep in the ocean depths (Lauro 15). They survive by reducing inorganic compounds, without the presence of oxygen, in the presence of great pressure. Microorganisms that are found in hot springs need to be able to thrive in harsh environments, as they are usually around 73 degrees (Costa 447). As said before, protein synthesis is often inhibited by high temperature or high pH, so the genetic differences of Thermoanaerobacter thermohydrosulfuricus and Caldanaerobacter subterraneus subspecies tengcongenesis and mesophilic Escherichia coli are notable when coding for exoproteins. The transformation was successful and Escherichia was able to produce these novel proteins, usually from bacteria that are completely anaerobic and use sulfur in their metabolic pathway in reduction-oxidation reactions.
Antranikian G. (2008) Industrial relevance of thermophiles and their enzymes. In: Rob et al. (eds) Thermophiles—biology and technology at high temperature. CRC Press Boca Raton. Pp 113-160.
Costa C. Kyle, Navarro B. Jason, Shock L. Everett (2009) Microbiology and geochemistry of great boiling and mud hot springs in the United States Great Basin. Extremophiles 13:447-459.
Lauro Frederico, Bartlett Douglas. (2007) Prokaryotic lifestyles in deep sea habitats. Extremophiles 12:15-25
Royter Marina, Schmidt M., C. Elend. (2009) Thermostable Lipases from the thermophilic anaerobic bacteria Thermoanaerobacter thermohydrosulfuricus SOL1 and Caldanaerobacter subterraneus subsp. Tengcongenesis. Extremophiles 13:769-783.





































